其它组分:
Cytochrome c (D18C7) Rabbit mAb 【子货号:#11940,包装:40 µl,,运保温度:–20°C】
Cox IV (4D11-B3-E8) Mouse mAb 【子货号:#11967,包装:40 µl,,运保温度:–20°C】
Smac/Diablo Mouse mAb 【子货号:#2954,包装:40 µl,,运保温度:–20°C】
Anti-rabbit IgG, HRP-linked Antibody【子货号: #7074,包装:100 µl,,运保温度:–20°C】
Anti-mouse IgG, HRP-linked Antibody 【子货号:#7076,包装:100 µl,,运保温度:–20°C】
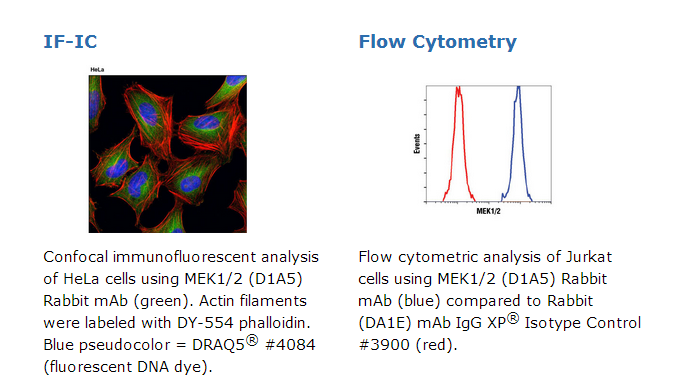
MEK1/2 (D1A5) Rabbit mAb 【子货号:#8727,包装:40 µl,,运保温度:–20°C】
Caspase-3 (8G10) Rabbit mAb 【子货号:#9665,包装:40 µl,,运保温度:–20°C】
HtrA2/Omi (D20A5) Rabbit mAb 【子货号:#9745,包装:40 µl,,运保温度:–20°C】
描述:
Caspase-3 (8G10) Rabbit mAb 兔单抗能够检测内源性的caspase-3 全长蛋白(35 kDa)和其Asp175处裂解(17/19 kDa)大片段。Cox IV (4D11-B3-E8) Mouse mAb 鼠单抗能够检测内源性的Cox IV总蛋白。Cytochrome c (D18C7) Rabbit mAb 兔单抗能够检测内源性的细胞色素C总蛋白。HtrA2/Omi (D20A5) Rabbit mAb 兔单抗能够检测内源性的HtrA2/Omi总蛋白,并且不与 HtrA1发生交叉反应。MEK1/2 (D1A5) Rabbit mAb 兔单抗能够检测内源性的MEK1和MEK2总蛋白,推测该抗体能与许多物种中的同源物发生交叉反应。Smac/Diablo Mouse mAb 鼠单抗能够检测内源性的加工和未加工的人Smac/Diablo蛋白,该抗体与小鼠和大鼠Smac/Diablo蛋白有弱的反应。
试剂盒中每种抗体均用Cell Fractionation Kit #9038进行了验证。这些单克隆抗体分别是由大肠杆菌提取的人源Smac/DIABLO蛋白全长,或用人源caspas-3 Asp175氨基末端、人源COX IV蛋白近C末端周围残基、人源细胞色素c蛋白Pro72周围残基、人HtrA2/Omi蛋白Phe341周围残基肽段或人源MEK1蛋白Ala220周围残基合成相对应的肽段免疫动物而制备的。The Apoptotic Release Antibody Sampler Kit 提供了一种经济有效的方法,检测凋亡应激下线粒体中释放的靶蛋白。试剂盒包括足够的一抗,足以完成四次迷你免疫印迹实验。细胞凋亡是一个被调控的生理过程,导致细胞死亡。Caspases,是一个半胱氨酸蛋白酶家族,是细胞凋亡的中心调控因子。引发剂 caspases(包括8、9、10和12)被紧密耦合到促凋亡信号。一旦被激活,这些 caspases 切割并激活下游的效应 caspases(包括3、6和7),它们反过来切割细胞骨架和核蛋白质,如PARP、α-胞衬蛋白、DFF和 lamin A,并诱导细胞凋亡(1)。细胞色素C是一种保守的电子传递蛋白,作为呼吸链的组分存在于于线粒体内膜中(2)。受到凋亡刺激信号后,细胞色素C从线粒体释放,并与procaspase-9 (47 kDa)/Apaf 1交联。此交联复合物促使caspase-9从没有活性的酶原转变成活化状态(3)。这一过程将进一步激发caspase-3的活化最终导致凋亡(4)。Smac/Diablo是一个21 kDa的哺乳动物线粒体蛋白,是细胞凋亡过程中的调控组件(5,6)。线粒体应激后,从线粒体释放的Smac/Diablo与半胱天冬酶竞争性结合IAPs(凋亡抑制蛋白)(5,6)。Smac/Diablo与IAP的相互作用解除了IAP对半胱天冬酶的抑制效果(7,8)。高温环境需要A2蛋白(HtrA2)/Omi是一种丝氨酸蛋白酶,与大肠杆菌的HtrA蛋白(DegP) 分子同源。研究认为,其与细胞凋亡及错误折叠蛋白质的降解引起的应激相关(9)。HtrA2产生时是50 kDa的酶原,之后被裂解以成36kDa的成熟蛋白,暴露出一个氨基末端的基序(AVPS),类似IAP抑制子Smac/Diablo(10-14)。与SMAC一样,HtrA2和IAP家族成员如XIAP之间的互作会拮抗抑制caspase活性,避免细胞凋亡(10-14)。Caspase-3 (CPP-32, Apoptain, Yama, SCA-1)是一个凋亡的关键执行者,因为它对许多关键的蛋白起着部分或全部的裂解作用,比如核酶多聚(ADP-ribose)聚合酶(PARP)(1)。caspase-3的活化需要经过其未激活的的酶原转变成活化的 p17 和 p12 片段的蛋白水解过程。caspase-3的剪切需要天门冬氨酸位于p1位点(16)。细胞色素c氧化酶(COX)是一个异寡聚酶,由13个亚基组成,定位于线粒体内膜(17-19)。它是呼吸链末端的酶复合体,催化分子氧还原为水,耦合跨线粒体内膜的质子转移,驱动ATP合成。3个大亚基形成催化核心,由线粒体DNA编码,而其他小的亚基,包括COX IV,由核基因编码。在细胞研究试验中,COX IV抗体可以作为有效的线粒体内参。MEK1、MEK2,也被称为MAPK或Erk激酶,它们是双特异性蛋白激酶,在丝裂原活化蛋白激酶级联反应中发挥作用,控制细胞生长和分化(20-22)。MEK1、MEK2的活化是通过Raf样分子对两个丝氨酸(217 /221位点)的磷酸化而实现的,这两个丝氨酸位点位于亚结构域VIII的活化环内。多种生长因子和细胞因子以及膜去极化和钙内流都能激活MEK1/2(20-23)。MEK蛋白磷酸化激酶亚结构域VIII活化环内的丝氨酸和苏氨酸残基后,就能激活p44和p42 MAP激酶。在基于细胞试验的研究中,MEK1/2 (D1A5) Rabbit mAb兔单抗能够作为一个有效的细胞质内参。
原厂资料:
Specificity / Sensitivity
Caspase-3 (8G10) Rabbit mAb recognizes endogenous levels of full-length (35 kDa) and large fragment (17/19 kDa) caspase-3 protein resulting from cleavage at Asp175. Cox IV (4D11-B3-E8) Mouse mAb recognizes endogenous levels of total Cox IV protein. Cytochrome c (D18C7) Rabbit mAb recognizes endogenous levels of total cytochrome c protein. HtrA2/Omi (D20A5) Rabbit mAb recognizes endogenous levels of total HtrA2/Omi protein and does not cross-react with HtrA1. MEK1/2 (D1A5) Rabbit mAb recognizes endogenous levels of total MEK1 and MEK2 proteins. This antibody is predicted to cross-react with MEK1/MEK2 orthologs in a variety of species. Smac/Diablo Mouse mAb recognizes endogenous levels of processed and unprocessed human Smac/Diablo protein and reacts weakly with mouse and rat Smac/Diablo.
Each antibody in the kit has been validated using the Cell Fractionation Kit #9038.
Source / Purification
Monoclonal antibodies are produced by immunizing animals with a full-length peptide corresponding to human Smac/Diablo protein raised in E. coli, or with synthetic peptides corresponding to amino-terminal residues adjacent to Asp175 of human caspase-3 protein, residues near the carboxy terminus of human Cox IV protein, residues surrounding Pro72 of human cytochrome c protein, residues surrounding Phe341 of human HtrA2/Omi protein, or residues surrounding Ala220 of human MEK1 protein.
Description
The Apoptotic Release Antibody Sampler Kit provides an economical means to evaluate targets that are released from the mitochondria with apoptotic stimuli. The kit contains enough primary antibody to perform four western blots per primary antibody.
Background
Apoptosis is a regulated physiological process leading to cell death. Caspases, a family of cysteine acid proteases, are central regulators of apoptosis. Initiator caspases (including 8, 9, 10, and 12) are closely coupled to proapoptotic signals. Once activated, these caspases cleave and activate downstream effector caspases (including 3, 6, and 7), which in turn cleave cytoskeletal and nuclear proteins like PARP, α-fodrin, DFF, and lamin A, and induce apoptosis (1).
Cytochrome c is a well conserved electron-transport protein and is part of the respiratory chain localized to the mitochondrial intermembrane space (2). Upon apoptotic stimulation, cytochrome c released from mitochondria associates with procaspase-9 (47 kDa)/Apaf-1. This complex processes caspase-9 from inactive proenzyme to its active form (3). This event further triggers caspase-3 activation and eventually leads to apoptosis (4).
Smac/Diablo is a 21 kDa mammalian mitochondrial protein that functions as a regulatory component during apoptosis (5,6). Upon mitochondrial stress, Smac/Diablo is released from mitochondria and competes with caspases for binding of inhibitor of apoptosis proteins (IAPs) (5,6). The interaction of Smac/Diablo with IAPs relieves the inhibitory effect of the IAPs on caspases (7,8).
High temperature requirement protein A2 (HtrA2)/Omi is a serine protease with homology to the E. coli HtrA protein (DegP) and is thought to be involved in apoptosis and stress-induced degradation of misfolded proteins (9). HtrA2 is produced as a 50 kDa zymogen that is cleaved to generate a 36 kDa mature protein that exposes an amino terminal motif (AVPS) resembling that of the IAP inhibitor Smac/Diablo (10-14). Like Smac, interaction between HtrA2 and IAP family members, such as XIAP, antagonizes their inhibition of caspase activity and protection from apoptosis (10-14).
Caspase-3 (CPP-32, Apoptain, Yama, SCA-1) is a critical executioner of apoptosis, as it is either partially or totally responsible for the proteolytic cleavage of many key proteins, such as the nuclear enzyme poly (ADP-ribose) polymerase (PARP) (15). Activation of caspase-3 requires proteolytic processing of its inactive zymogen into activated p17 and p12 fragments. Cleavage of caspase-3 requires the aspartic acid residue at the P1 position (16).
Cytochrome c oxidase (COX) is a hetero-oligomeric enzyme consisting of 13 subunits localized to the inner mitochondrial membrane (17-19). It is the terminal enzyme complex in the respiratory chain, catalyzing the reduction of molecular oxygen to water coupled to the translocation of protons across the mitochondrial inner membrane to drive ATP synthesis. The 3 largest subunits forming the catalytic core are encoded by mitochondrial DNA, while the other smaller subunits, including COX IV, are nuclear-encoded. The Cox IV (4D11-B3-E8) Mouse mAb can be used effectively as a mitochondrial loading control in cell-based research assays.
MEK1 and MEK2, also called MAPK or Erk kinases, are dual-specificity protein kinases that function in a mitogen activated protein kinase cascade controlling cell growth and differentiation (20-22). Activation of MEK1 and MEK2 occurs through phosphorylation of two serine residues at positions 217 and 221, located in the activation loop of subdomain VIII, by Raf-like molecules. MEK1/2 is activated by a wide variety of growth factors and cytokines, as well as by membrane depolarization and calcium influx (20-23). MEK activates p44 and p42 MAP kinase by phosphorylating both threonine and tyrosine residues at sites located within the activation loop of kinase subdomain VIII. The MEK1/2 (D1A5) Rabbit mAb can be used effectively as a cytoplasmic loading control in cell-based research assays.



 京公网安备11010802025653 版权所有:北京逸优科技有限公司
京公网安备11010802025653 版权所有:北京逸优科技有限公司
